Article Text

Statistics from Altmetric.com
The HIV/AIDS pandemic has so far caused about 35 million deaths, while 34 million individuals currently live with HIV.1 ,2 Even if this will have no impact on the course of the epidemic, understanding the factors that allowed the successful emergence of HIV-1 is important, first as a moral obligation towards the victims, but also to draw lessons that could ultimately help mankind avoid facing similar threats in the future.
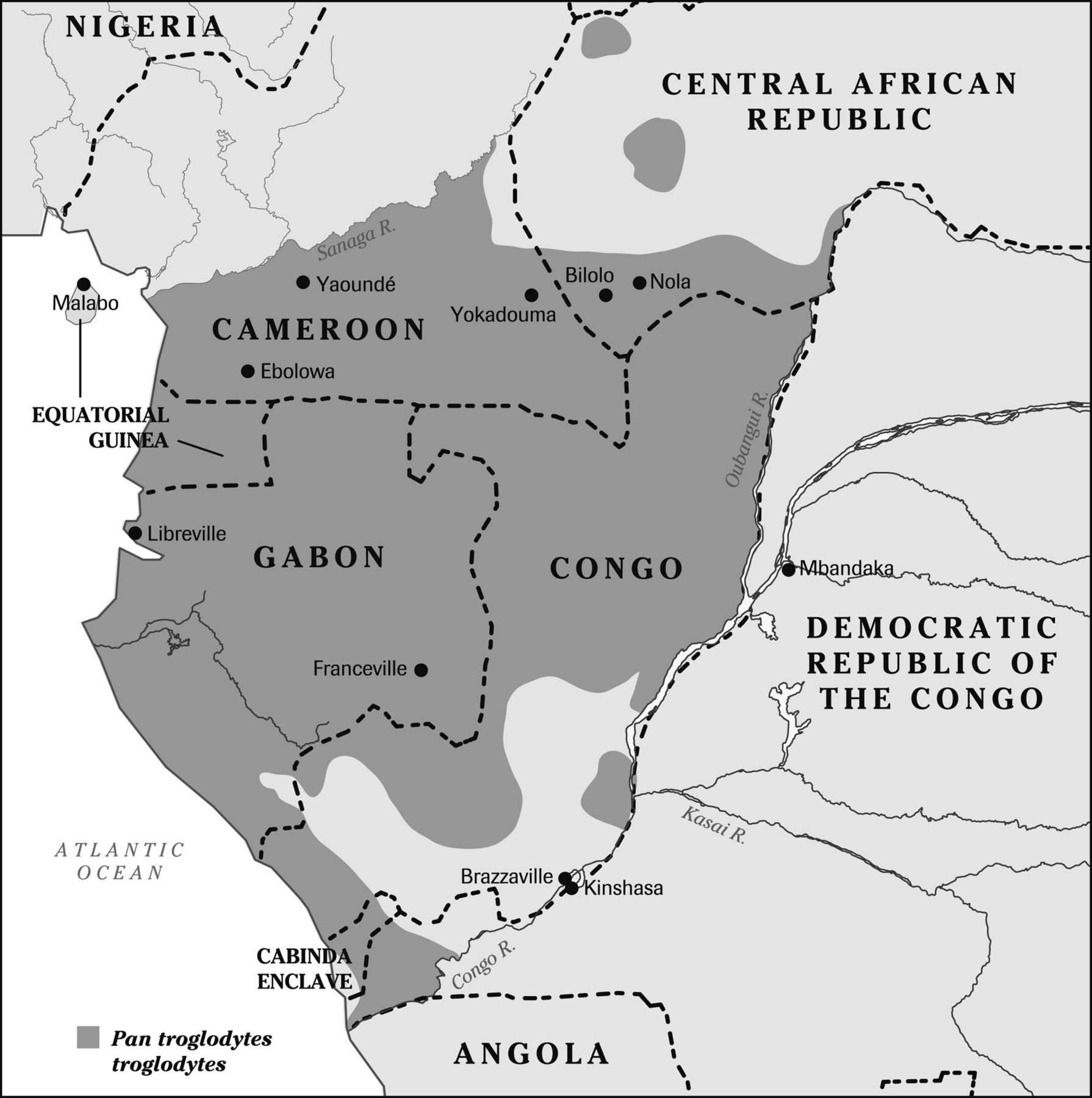
Over the last 12 years, much progress has been made in unravelling the complex chain of events that led to the worst pandemic of modern times. The source of HIV-1 group M (‘M’ for main) is the Pan troglodytes troglodytes chimpanzee of central Africa3 ,4 which inhabits southern Cameroon, Gabon, Equatorial Guinea, the Congo–Brazzaville, the southwest of the Central African Republic, the Cabinda enclave of Angola and a small part of the Democratic Republic of Congo north of the Congo river (figure 1). The true ‘Patient Zero’, the one who started the pandemic, must have lived somewhere within this territory. Sequencing evidence suggests that this may indeed have been in southeast Cameroon or the adjacent areas of the Central African Republic and Congo–Brazzaville. But after some local transmission to start with, HIV-1 eventually managed to flourish and diversify further down the Congo River.
{kind=link}
Map of central Africa showing the distribution of the Pan troglodytes troglodytes source of HIV-1.
Through sophisticated molecular clocks, it was estimated that the original cross-species transmission of HIV-1 group M, from chimp to man, occurred during the first three decades of the 20th century. By exclusion of other hypotheses, this initial event probably occurred through the manipulation of chimpanzee meat by a hunter or his wife, who manipulated the ape's carcass in order to cook it.1 Three other ‘groups’ of HIV-1 (groups O, N and P) have been identified. They managed to infect a much more limited number of humans (≈10 000 for O, 14 for N, 2 for P, most of them Cameroonians), and are thought to have resulted from different cross-species events, each chain of transmission starting with a single infected person. If four different cross-species events, one for each group, could be recognised decades later, it is likely that other hunters or hunters’ wives got infected prior to the 20th century, resulting in epidemiological dead ends. The hunter infected his wife, both died of AIDS in their village, and that was the end of it. But then, why did HIV-1 group M become so successful in the 20th century, eventually infecting 69 million people worldwide?
Some of the epidemiological success of group M, as opposed to the limited spread of groups O, N and P, may have been driven by biological characteristics of the virus. For instance, tetherin, a human protein that normally protects us from zoonotic viruses, is less effective against group M than against the other groups.5 But this is unlikely to explain the full story, unless one postulates that no hunter had ever been infected earlier than the 20th century with simian ancestors of group M. It seems more plausible that two factors intimately linked to the European colonisation of central Africa facilitated the emergence of HIV-1: urbanisation and healthcare.1
The French and Belgian colonisers created small cities where, for several reasons, the number of unmarried men far exceeded the number of unmarried women. Naturally, this imbalance led to sex trade, but for a long time prostitution in central Africa, especially in Léopoldville and Brazzaville, was of a low-risk type.1 Les femmes libres (‘free women’) had three or four regular clients, each visiting the woman once a week and receiving diversified services which included sexual intercourse. This degree of concomitant sexual partnerships was good enough for the virus to persist and slowly expand, but not for an exponential amplification. The exponential amplification of HIV-1 through heterosexual transmission, for instance, what occurred in Nairobi in the mid-1980s, requires high-risk high-volume prostitution, in which women have sex with a thousand different men each year.6
Early in the process, another factor allowed HIV-1 to expand. Starting in Cameroon and French Equatorial Africa during World War I, quickly imitated by the Belgian Congo, colonial health authorities implemented ambitious programmes aiming to control selected tropical diseases. Initially, sleeping sickness was targeted, but syphilis, yaws and leprosy soon followed.1 Twice a year, dedicated mobile teams examined the whole population of each and every village. Participation was compulsory. Those found to have one of these diseases were treated locally by nurses who stayed back after the rest of the team had departed. Antimicrobial drugs were not very effective then; to maximise their clinical impacts, many had to be injected intravenously, generally once a week for 3–15 consecutive weeks, depending on the diagnosis. The existence of viruses was just beginning to be hypothesized, and nobody anticipated that bloodborne viruses could be transmitted by injections. Needles and syringes were used repeatedly on any given day with improper, if any, sterilisation. In many communities of southern Cameroon, 50% of some birth cohorts were ultimately infected with the Hepatitis C virus (HCV), indicating the massive iatrogenic spread of at least one bloodborne virus.7 In one of these towns called Ebolowa, the intravenous treatment of malaria was the principal route of transmission of HCV among elderly people.8 Around Nola in the Central African Republic, the worst ever focus of sleeping sickness during the colonial era, located within the area where ‘Patient Zero’ probably lived, HCV was transmitted through the treatment of this parasitic disease before 1950, while Human T-cell lymphotropic virus (HTLV-1), another retrovirus with a chimpanzee reservoir, was spread through the injections of pentamidine given twice a year to the whole population for the prevention of trypanosomiasis.9 Patients treated for sleeping sickness in Nola in the 1940s experienced a staggering excess mortality, potentially compatible with the iatrogenic dissemination of HIV-1.9 In another part of the continent, an epidemiological study of elderly people in Guinea–Bissau revealed that HIV-2 had been transmitted through the treatment of sleeping sickness but also of tuberculosis (with long courses of intramuscular streptomycin).10 Thus, in three different countries, different colonial-era tropical diseases therapeutic interventions resulted in the transmission of three different bloodborne viruses. By contrast with the highly lethal HIV-1, these viruses are compatible with a prolonged survival, enabling such associations to be documented retrospectively. Immunisations probably did not play much of a role, because the vaccines then available against smallpox and yellow fever were administered intradermally, a rather ineffective route for transmitting bloodborne viruses.
The second and most crucial stage of the pandemic took place in the Léopoldville–Brazzaville binational conurbation, the terminus for all river transportation on the huge Congo basin (some of whose tributaries drain southeast Cameroon). Not only were the two oldest isolates of HIV-1 recuperated from samples obtained in Léopoldville in 1959 and 1960, but molecular studies revealed that the Léopoldville–Brazzaville area harbours, by far, the highest genetic diversity of HIV-1 group M in the world.11–15 All nine ‘subtypes’ of HIV-1 group M have been found there, as well as many recombinants.14 By contrast with groups, subtypes represent the evolution of HIV-1 within humans, the accumulation of replication errors. Their diversity reflects not only the duration of the presence of HIV-1 in a given place but also the efficacy of its transmission (if more persons get infected, more copies of the virus are produced each day and more replication errors intervene).
Once the virus reached the Belgian Congo's capital, the number of infected persons very slowly increased during the first half of the century, followed by an exponential amplification starting in the early 1950s.12 The semiprostitution practiced at the time by free women could not possibly account for this steep rise in the number of HIV-infected persons, but what about parenteral transmission? In Léopoldville, tropical diseases were uncommon as the African population was regularly screened and treated so as to protect the Belgians. But the local sexually transmitted diseases (STD) clinic may have represented a perfect storm: it treated free women who had to show up monthly to be in line with the law, and migrants were also compelled to be screened. Staggering numbers of injections were administered in this small facility (peaking at 154 572 in 1953), mostly for the treatment of presumed syphilis.16 An irony is that, in retrospect, the overwhelming majority of these patients never had syphilis: their non-treponemal serology was positive because of yaws acquired during childhood.16 In 1953, an outbreak of ‘inoculation hepatitis’ (presumably, Hepatitis B) developed among patients treated at the STD clinic where injection devices were merely rinsed between patients.17 One can imagine that if the Hepatitis B virus thrived within this facility, HIV-1 might also have followed the same path of iatrogenic transmission to free women, most of whom were having sex with 3–4 regular clients.
To summarise, it seems likely that between 1900 and 1930, a ‘Patient Zero’ acquired Simian immunodeficiency virus (SIV)cpz through manipulation of chimpanzee meat somewhere near the southeast corner of Cameroon. At the very moment of this cross-species event, SIVcpz became HIV-1: the same virus, with the same genome, in a new host. Local transmission ensued, probably mostly through the intravenous treatment of sleeping sickness and other tropical diseases. A first critical mass of HIV-infected persons was reached, maybe a few hundreds, so that it became unavoidable that a few of them would travel to the local metropolis of Léopoldville–Brazzaville, where sexual and/or parenteral transmission allowed the virus to persist. In the early 1950s, exponential amplification occurred mainly through parenteral transmission, followed in the 1960s by amplification via the heterosexual route, which became possible when the face of prostitution in the city changed abruptly, as the upheavals provoked by the Congo's botched decolonisation led to mass migrations into Léopoldville and severe poverty. Men who could no longer regularly support a free woman started buying sex on a fee-for-service basis.18 Beginning in the 1960s and continuing into the 1970s, the virus was exported out of Léopoldville to other African countries, but also to the USA via Haiti, probably through one of the 4500 Haitian technical assistants who worked in the Congo during this turbulent period.1 From Haiti, the virus was quickly re-exported to the USA, presumably through the sexual tourism of American gay men in the 1970s. The USA then seeded many other locations, mostly in Western Europe and Latin America. Within Africa, out of their Léopoldville bridgehead, different subtypes disseminated along various routes. For instance, subtype C eventually reached Elisabethville (now Lubumbashi), from where it crossed the border into Zambia, eventually reaching South Africa by the way of Malawi and Zimbabwe.1 Léopoldville truly had been Ground Zero for HIV-1.
The above narrative represents what seems to me the most likely scenario, but of course it will never be possible to reconstruct perfectly the complex history of the HIV-1 pandemic, and there is room for opinions to differ. While many pieces of the puzzle have now been correctly positioned, a few more remain to be worked out. A more precise timeline might be delineated, should it prove possible to recuperate additional ancient isolates of HIV-1. Hopefully, clever molecular biologists will eventually explain to us why the SIVcpz of another chimpanzee, Pan troglodytes schweinfurthii, which inhabits the Democratic Republic of the Congo (DRC), did not manage to infect humans, despite opportunities which must have been similar to those of P t troglodytes.20 And why did HIV-1, subtype B, disseminate successfully in North America, the Caribbean and Western Europe while remaining so rare throughout Africa?1
So which lessons can be drawn from this story? An obvious one is that medical interventions, or more generally scientific initiatives, can have disastrous and unpredictable long-term consequences: more prudence and humility from scientists would be useful for the future of mankind. For instance, one wonders whether manipulating the genome of the H5N1 influenza virus to enhance its transmissibility between humans is really a good idea. A second lesson is how our world has truly become a ‘global village’, to use the words of McLuhan: starting with a hunter in a remote village of Central Africa, the aetiological agent of AIDS managed to infect and kill millions in all continents. It is clearly now in the own interest of industrialised societies to invest resources into the control of infectious diseases of the Third World, through the Global Fund and other initiatives. Hopefully, we can do it this time without spreading a novel infectious agent.
References
Footnotes
-
Funding None.
-
Competing interests None.
-
Provenance and peer review Commissioned; externally peer reviewed.