Article Text

Abstract
Objective: To assess claims that genes are a major determinant of social class.
Design: Using genetic epidemiological principles, five claims on the role of genes in determining social class are examined: (1) traits that run in families are usually inherited; (2) complex traits can be explained by alleles at a single gene locus; (3) complex traits are transmitted intact from one generation to the next; (4) natural selection explains social advantage. (5) Heritability estimates provide a valid estimate of the importance of genes in explaining complex human traits or behaviour.
Results: (1) Traits that run in families can result from environmental exposures that differ by social class. (2) The protein encoded by any single gene has too narrow a range of biological activity to explain traits as complex as social status. (3) Because alleles at different gene loci are transmitted independently, genetic inheritance cannot explain why offspring display the same complex traits as their parents. (4) The propagation of mutations that might result in a selective advantage takes much longer than the time for which any social class has achieved or maintained dominance. (5) Heritability measures are accurate only when environment is maintained constant. This is impossible in evaluating human traits.
Conclusions: The roots of social class differences do not lie in our genes. Consequently, genetics cannot be used as a justification for maintaining a ruling class, limiting procreation among the poor, or minimising social support programmes.
- genetics
- social class
- heredity
- environment
- equality
Statistics from Altmetric.com
Heredity has been a justification for power from ancient times to the present. Royal lineages were maintained by claiming that the “divine right” of kings was inherited. Darwin's theory of natural selection provided a biological basis for justifying social status. Coining the term “eugenics,” Darwin's cousin, Francis Galton, maintained that those with “good” traits, which he considered to be those of the professional classes in England,1 (9, 76) had a duty to have more children. The turn of the 20th century brought the rediscovery of the work of Gregor Mendel and the notion of the gene as the unit of inheritance. At about the same time, procedures for both male and female sterilisation were developed, offering a means of curtailing the reproduction of those with “undesirable” traits.2 In the United States, Charles Davenport and his colleagues began to collect pedigrees displaying a wide range of physical and social traits. They argued that inherited traits explained class differences. Criminality and mental deficiency rather than being the effect of poverty were the cause.1,3 (It is interesting to note that some of the eugenic sterilisation laws passed in the United States excluded white collar crimes such as embezzlement, a point that led the Supreme Court to declare the Oklahoma law unconstitutional by denying the equal protection guaranteed by the 14th Amendment to the Constitution.4) To both Galton and Davenport, the observation that “like begets like” was a strong argument for the importance of genetics. This belief has persisted. A front page story in the New York Times in 1992 attributed the finding that more than half of prisoners had relatives who were also convicted criminals at least in part to inherited genetic factors.5
The mapping of the human genome accelerated searches for associations between complex diseases or behavioural traits and specific regions of chromosomes that, presumably, contained the genes whose variation accounted for the association. At this stage, too, some scientists maintained that class differences could be explained in genetic terms. “No group would benefit more from the genome project than the homeless,” Daniel Koshland, then editor of Science said,6 because diseases like depression and schizophrenia led them into poverty and, consequently, homelessness.
The sequencing of the human genome 7,8 facilitates direct identification of the genetic variations that might be associated with complex behavioural and social traits. The ability to identify human genes has encouraged the belief, that, as James Watson, told a reporter, “our fate is in our genes,”9 and it is only a matter of time before the genetic basis of the vast majority of human experience is discovered.
As the 20th century ended, it became increasingly apparent that many traits could not be explained on the basis of variations in genes at single loci. Earlier, the heritability statistic was devised to partition the variance in complex traits between genetic and environmental components. Claiming high heritability for traits that correlated with social class, Richard Herrnstein 10 argued that people will sort themselves into classes because of genetic differences and these differences will be perpetuated in succeeding generations of the various classes.
In this paper, I will examine the claims made for an association between genes and social class. Using principles of genetics and how genes function, I will show that genetics cannot explain the perpetuation of complex traits in one class in succeeding generations and why alleles at a single gene locus fail to account for most complex traits. If more geneticists, molecular biologists, and epidemiologists, as well as social scientists, bioethicists, and philosophers, grasped these principles, there might be fewer exaggerated claims for the role of genes. Inherited genetic factors cannot provide the explanation of social organisation.
THE USE OF PEDIGREES AND FAMILY HISTORY TO BOLSTER A GENETIC BASIS FOR CLASS DIFFERENCES
In the early 20th century, the Eugenics Record Office (ERO) at Cold Spring Harbor, NY under the direction of Charles Davenport, set out to collect pedigrees of various traits. Traits that ran in families, particularly if they were consistent with Mendel's laws of inheritance, were assumed to be inherited. Pellagra was a disease of poor people. Davenport's hypothesis, “that in the pellagra reaction there is a hereditary factor” 11 (7) would, if proven, establish a genetic factor that was more prevalent among the poor than among the wealthy and could, therefore, explain poverty. The factor might be “mental insufficiency,” which Davenport and other eugenicists perceived to be much more prevalent among the poor than the rich:
“ (T)he mentally insufficient are, on the whole, less likely to appreciate the importance of sanitary surroundings and less able to avail themselves of them...(P)ersons who are mentally developed are, on the whole, more likely to care for their bodies and keep themselves in good condition than are the mentally deficient or unstable. Other things being equal, pellagra is more likely to make headway in `Nam Hollow' than in the cottages on the cliffs at Newport.”11 (The Nams were an impoverished family of alleged mental defectives studied by the ERO. Newport was the wealthy Rhode Island resort community.)
The thought never seems to have dawned on Davenport (or on Koshland almost 75 years later) that poverty might itself interfere with people's ability “to keep themselves in good condition.”
Confirming Davenport's hypothesis proved difficult. In analysing data from extensive field studies, Dr Elizabeth Muncey, Davenport's colleague at the ERO, found many features that did not fit with an inherited factor: (1) Fewer children of pellagrous parents had pellagra than would be expected if the disease was attributable to a single allele accounting for an autosomal dominant trait. (If pellagra was an autosomal dominant trait, then one half of children in which one parent had pellagra would also have it. Instead, only 16% had it.) (2) Almost as many pellagrous children were born to couples in which neither parent was pellagrous as to couples in whom one or both were. An autosomal recessive trait could account for unaffected couples having children with pellagra, but then all children born to parents who were both pellagrous should also be affected. Only 18% were. In addition, the observed proportion of affected children born to unaffected parents exceeded the one quarter expected for an autosomal recessive trait. This finding is also inconsistent with alleles at more than one gene locus being necessary for pellagra unless the loci were tightly linked, which is unlikely. (3) Many more daughters than sons were affected regardless of which parent (if either) was affected. (4) An insignificant number of families had three affected generations. This also argued against a dominant mode of inheritance. Finally, (4) cases among siblings arose in close temporal proximity. Muncey concluded, “the data collected shows no evidence of direct heredity, There may, however, be an hereditary predisposition to the disease in those families in which chronic gastro-intestinal symptoms have existed for several generations .”12 No data in the paper support this contention.
In the accompanying paper, Davenport contradicts at least one of Muncey's findings. Whereas she reported that only 18% of children of parents who were both pellagrous were also pellagrous, Davenport claims,
“When both parents are susceptible to the disease, at least 40 percent., probably not far from 50 per cent., of their children are susceptible; an enormous rate of incidence in a disease that affects less than 1 per cent. of the population on the average.”11
(Davenport may mean something other than overt disease when he speaks of “susceptible,” but he never makes it clear.) Like Muncey he concludes that the high incidence among siblings
“is doubtless due to infection, (but) it is also doubtless due to susceptibility, for right among the affected children grow up brothers and sisters who have never shown symptoms of pellagra.”
Davenport added another argument for the role of heredity in pellagra, racial differences:
“This constitution of the organism is a racial, that is, hereditary factor. And if it appears that certain races or blood lines react in the pellagra families in a specific and differential fashion, that will go far to prove the presence of a hereditary factor in pellagra.”
Citing observations of others “that colored (sic) persons ...are less subject on the whole to the disease than white persons,” Davenport concludes, “We thus have strong evidence of a racial difference, and that is synonymous with a hereditary difference .”11 Davenport does cite evidence of pellagra in “negroes” and concedes, “it is difficult to draw correct inferences as to susceptibility from the apparent difference in morbidity between the negro and white population.”11
On even more slender evidence, Davenport concludes that the different clinical manifestations of pellagra (gastrointestinal, skin, nervous system) are attributable to “biotypes that differ in the specific resistance or susceptibility of their different organs.” By biotypes he means different alleles or other genetic factors. Davenport concludes his paper with the following sentence:
“These family differences have all the characteristics of biotypes or blood lines, and afford the best proof that there is, indeed, a hereditary factor in pellagra .”11
Much more telling than Muncey's failure to find “direct evidence of heredity” for pellagra, were the studies of Joseph Goldberger, published in 1915,13 before Davenport even submitted his paper on 9 March 1916. There can be no doubt that Davenport was aware of Goldberger's work as the pellagra Commission with which he collaborated (and wrote part of the final report) was aware of it and several members withdrew afer Goldberger's work was published .14
Goldberger drew quite a different inference from the fact that pellagra was concentrated among the poor:
“One of the outstanding features of the epidemiology of pellagra is the striking relation of the disease to poverty...(I)t seemed permissible to assume, on the one hand, that the diet of the poor, that is, of those who as a class are the principal sufferers of the disease, is, for some reason, pellagra-producing, and, on the other, that the diet of the well-to-do who, as a class are practically exempt, is for some reason, pellagra-preventing.”15
Goldberger showed that pellagra could be produced experimentally by feeding volunteers diets devoid of fresh animal protein and legumes.13,15 Moreover, the disease could be successfully treated by adding these foods to the diet of pellagrous patients while keeping other factors constant.15 If an inherited factor played a part it would have to be present in a large proportion of the population; 6 of 11 men in Goldberger's study developed pellagra within six months of being placed on a deficient diet. This contrasts with Davenport's estimate that only 1% of the population develops pellagra. Moreover, Goldberger found pellagra among “colored” patients. The finding that dietary deficiency accounts for pellagra also explains Muncey's findings of more pellagra among women than men. As the breadwinners or potential breadwinners, men were given better diets than women in many households.
We can only speculate as to why Davenport persisted in his belief that a hereditary factor was important in pellagra. Chase offers the following explanation:
“For let it once be revealed that pellagra, like hookworm, was not in the genes but in the debilitating environment of southern poverty, then the entire eugenic-dysgenic basis [which Davenport espoused] for characterizing America's Nordic [white] poor as being genetically ineducable, unemployable, and unfit for decent housing went careening down the drain.”14 (204).
Writing in more general terms about the motivations of eugenicists and those who supported them, Garland Allen concludes:
“By suggesting a genetic cause for persistent or recurrent social dilemmas, hereditarian theories suggest that the victims, not the social system, are the cause of their own problems...(T)he solution is not to be found in political and social change but, instead, in change in the genetic makeup of the population.” 3
Although the antecedents of pellagra have now been shown not to be genetic, except in those with a rare recessive disorder (Hartnup's disease 16), poverty and social class status continue to be attributed to genes. Before exploring this association further, we need to examine what genes do.
ALLELES AT SINGLE GENE LOCI DO NOT DETERMINE COMPLEX TRAITS
Alleles at single gene loci do not encode for complex traits, including those that might correlate with social status, but for a protein (or subunit thereof), which may, depending on the gene locus, function as an enzyme, cell receptor or channel, hormone, or transporter of other molecules. At additional gene loci, alleles regulate the transcription of other genes. If a person inherits an allele with a deleterious mutation, complex consequences may result, such as mental retardation. For instance, infants who inherit defective alleles for the enzyme phenylalanine hydroxylase from their mother and father will have phenylketonuria (PKU), which, in the absence of early treatment, almost invariably results in mental retardation. Particular genotypes of several dozen other genes will also result in mental retardation. Can we say, therefore, that any one of these genes when normal is the gene for intelligence? Although each of them may contribute to intelligence, even when all of the wild-type (normal) genotypes are present, there is no assurance of normal intelligence. A defect in another gene whose function is yet unknown could contribute to retardation. For the same reason, although the presence of abnormal alleles at any one of several different gene loci results in short stature, normal function of all of these genes will increase the probability of normal stature but not assure it. (It is possible that genetically engineered changes in a gene could improve its enzymatic or other function that might enhance a trait. The degree of enhancement would depend on the genetic and environmental background. The enhancement if it did occur would not be transmitted to the next generation unless the change was introduced into the person's germ cell, which no scientist has yet attempted for both ethical and technical reasons.)
Environmental factors will interact with genes and gene products (for example, proteins). Some alleles that encode for glucose 6-phosphate dehydrogenase result in haemolytic anaemia, but only when the people who possess them are exposed to certain drugs or foods.17 Among women with BRCA1 or BRCA2 alleles that increase susceptibility to breast cancer, those who were born before 1940 have a risk of breast cancer of 38% by age 55 whereas for women born after 1940 the risk is 69% (p<0.01).18 This cohort effect may be attributable to a difference in environmental exposures, perhaps oestrogen-like compounds to which the younger cohort have had greater exposure through oral contraceptives or higher concentrations of oestrogen-like substances in dairy, meat, and poultry products. Behavioural differences could also be an explanation.
In many cases environmental effects will swamp out genetic differences, just as a deficient diet was sufficient to explain the vast majority of cases of pellagra without having to invoke a genetic susceptibility. In the United States the proportion of neurobehavioural deficits attributable to increased lead levels exceeds that attributable to genetic and chromosomal abnormalities. One writer concluded, “Lead was certainly the most widespread threat to child health in America” in the 1980s.19 As nutrition improves in a country, average height increases from one generation to the next; this is too rapid to attribute to a change in the genes.
Thus for complex traits that might be correlated with social class, such as leadership capability, intelligence, entrepreneurship, aggressivity, or cunning, the presence of certain (almost entirely undefined) alleles at many gene loci as well as environmental factors play a part. Moreover, the alleles at different gene loci that make up this hypothetical constellation will usually be inherited independently of each other—that is, they reside on separate chromosomes or are far apart on the same chromosome. Thus if a single ruler happens to have a constellation that contributes to his ability to be a ruler, it is virtually impossible that he or she will pass it on intact to any of his sons or daughters. Moreover, different constellations of alleles at multiple loci could result in the same trait.20 These constellations will not be exclusively possessed by those in the ruling class. This will become clear when we examine the fallacy that genetic factors explain why “like begets like.”
“LIKE BEGETS LIKE” CAN SELDOM BE EXPLAINED ON A GENETIC BASIS
As we have seen, Davenport's hypothesis of a hereditary factor in pellagra was based in part on the appearance of the disease in succeeding generations, or “like begets like.” He and some other eugenicists assumed that this phenomenon was explainable on a genetic basis. Over 50 years later, Herrnstein's argument for a genetic basis for class differences was also based on “like begets like.”
“...the tendency to be unemployed may run in the genes of a family about as certainly as bad teeth do now...
“The higher the heritability, the closer will human society approach a virtual caste system, with families sustaining their position on the social ladder from generation to generation...” 10
In other words, Herrnstein argued, if society removed all social barriers to full equality, inequality because of genetic inherited factors would become worse. (I will discuss problems with heritability measurements below.)
But genetics does not always or even often explain why like begets like. Galton himself recognised that parents who were outliers for quantitative traits that he thought were inherited (for example, height) tended to have children whose same traits were closer to the mean.1 (15). By 1916, the year Davenport and Muncey published their papers on pellagra, most geneticists were aware of the Hardy-Weinberg principle, which made it clear that the frequency of an autosomal recessive trait in one generation would depend not primarily on matings between affected homozygotes (like begets like) in the previous generation but on matings between unaffected heterozygotes.21 If one assumes, as many eugenicists in the early part of the 20th century did, that mental deficiency was an autosomal recessive trait attributable to a single homozygous genotype (for example, “aa”), and that it occurred in 1% of any generation, then 81 times as many “mental defectives” would be born to mentally normal parents with heterozygous genotypes (“aA”) as to parents who themselves were mentally defective with the genotype “aa”. (In this model, the frequency of the “a” allele=0.1. and “aa”=a2 =0.01. Assuming only two alleles exist, the frequency of the “A” allele = (1−a) = 0.9. The frequency of heterozygotes =2aA=0.18 and the frequency of matings between heterozygotes = (2aA) × (2aA) =0.0324. One quarter of the offspring of matings between heterozygotes would be affected homozygotes, aa=0.0081. The frequency of matings between affected homozygotes = a2 x a2 =0.01× 0.01= 0.0001 and all of their children would be affected homozygotes =0.0001. Therefore, the ratio of affected homozygotes born to heterozygotes to those born to affected homozygotes =0.0081/0001=81. (The calculation ignores matings between heterozygotes and affected homozygotes, which would account for 18% of all affected homozygotes and bring the total frequency of “aa” to 0.01. It is also based on the assumption of random mating.) This mathematical exercise also makes it clear that sterilising “mental defectives,” even if all were attributable to the same homozygous genotype, would be a very inefficient way of reducing the frequency of the genotype in succeeding generations, but that is what many eugenicists and some geneticists advocated.21 If there were many different homozygous genotypes that resulted in mental deficiency, then retardation would not result from matings of “mental defectives” with each other unless both had the same genotype, or from matings between heterozygotes unless both partners carried the same allele for mental deficiency.
Matings between persons who possess a “favourable” recessive trait would yield only a small proportion of those who possess the trait, but it would result in like begetting like 100% of the time. One could argue that while others will possess the trait, such matings could serve as the nucleus of a social class. Non-random mating between homozygotes for a “favourable” recessive trait (for example, arranged marriages or like seeking like) would, however, increase the likelihood of homozygosity for alleles at other gene loci that result in autosomal recessive diseases in the offspring. The “favourable” trait would not survive for very long if these diseases curtailed survival and reproduction.
“Like begets like” occurs for autosomal dominant traits: three quarters of the offspring of two heterozygotes who each possess one copy of an allele (“a”) that encodes for a dominant trait will possess at least one copy of the “a” allele and will also be affected (assuming that the one quarter of the offspring who are homozygous for the allele will survive and manifest the trait; this is likely for a “favourable” trait) (see table 1) Non-random mating between those with “favourable” dominant traits would also increase the probability that offspring would inherit alleles at other loci for “unfavourable” dominant and recessive traits, including disease.
Genotypes arising from matings between AaBb persons when only two alleles occur (Aa, Bb) at two independently segregating loci*
Upper class status is much too complex a “favourable” trait to be the result of alleles at a single gene locus. The more the number of independently segregating loci needed before the trait manifests, the smaller the proportion of offspring who will display the trait. This is illustrated in table 1, where the proportion of offspring who display a trait for which the presence of a single allele (“a”) at one locus suffices is compared to the proportion when the presence of single alleles at two loci (“a” and “b”) is needed. Because of independent segregation the probability that the ovum or sperm of a woman or man respectively who is “AaBb” being “ab” is equally likely as it being “Ab”, “aB”, or “AB”. Instead of three quarters of offspring of such matings being affected (if only a single gene dose of the a allele was needed), now only 9 of 16 (56%) will be affected (the proportion inheriting both “a” and “b” alleles). Like begets like less and less as more independently segregating loci are involved. Even if people of a given “caste” will choose to mate only with each other, Herrnstein's argument that this would perpetuate the trait in succeeding generations does not hold up: The majority of the people manifesting the trait (for example, “upper caste”) will be heterozygotes at each of the required loci and will bear children without the trait because they lack both of the necessary alleles, “a” and “b”. After a relatively small number of generations, the descendants of those possessing the “desirable” trait will possess all possible genotypes at the imputed loci with regression towards the most frequent genotypes. CC Li put this succinctly:
“The most important single phenomenon of the genetic model is that for any given class of parents, their offspring will be scattered into various classes; conversely for any given class of offspring, their parents come from various classes. Environmentalists sometimes misunderstand the implications of population genetics, thinking that heredity would imply `like class begets like class.' Probably, the opposite is true. Only very strong social and environmental forces can perpetuate an artificial class; heredity does not.”22
As data have accumulated on DNA as well as protein variation within and among populations it can now be shown that the amount of genetic difference increases as the geographical distance between populations increases23 (23). Considerable admixture among groups living in close proximity occurs over several generations. The upper class in one geographical region may be of greater genetic similarity to the lower class in that region than to the upper class in a remote region, particularly when the two regions were isolated from each other until recently.
THE ROLE OF MUTATION AND NATURAL SELECTION
Part of the emphasis given to the importance of genes, and to single genes in particular, is based on the recognition that mutations arise in genes and the belief that the increasing frequency of “advantageous” mutations in subsequent generations (selection) operates at the level of the gene. As Mayr has pointed out:
“...not genes but whole organisms–potentially reproducing individuals–are the unit of selection. This means that the effects of recombination and of gene regulation, as well as the capacity of developing phenotypes to respond to the environment, are as important for selection as is mutation, indeed quantitatively actually more important by several orders of magnitude.” 24 (588)
Key points
-
Seldom if ever are traits associated with social class explained by genetic differences. These traits are usually the result of inequalities, not the cause.
-
When complex traits are influenced by genes, many independently segregating alleles are usually involved; they will not be transmitted together from one generation to the next.
-
Even if mating were rigorously controlled (non-random) one class could not propagate itself on genetic grounds.
-
Heritability measurements exaggerate the genetic, compared with the environmental, contribution and provide no information about the number of genes involved or how they result in the trait.
-
Genetics should cease to be a subterfuge for explaining social differences.
It is highly unlikely that a mutation at a single gene locus will of itself be of sufficient benefit to convey to its first bearer and to all subsequent bearers an advantage in reproductive fitness that is not also dependent on the alleles present at other gene loci as well as on the environment. Moreover, the replacement of an old allele with a new one as the most frequent allele may not necessarily occur by natural selection but by random drift. In neither case, however, will the process occur rapidly over a few generations. As Luca Cavalli-Sforza writes:
“It is difficult to distinguish between diffusion of a selectively neutral gene...because of drift and the spread of an advantageous mutation by natural selection...(T)he selective advantages are usually modest, and it may take thousands or tens of thousands of generations to substitute an improved gene. In humans, one thousand generations would span 25,000 years. If a gene presents a strong selective advantage, it can be spread by natural selection in only a few hundreds, or few thousands, of years.”23 (45)
Even if we were to concede that a single allele (in either heterozygous or homozygous gene dosage) were to convey a social advantage, no ruling class has ever remained in power long enough to propagate that putative allele in subsequent generations.
THE USE OF HERITABILITY TO BOLSTER A GENETIC BASIS FOR CLASS DIFFERENCES
“Heritability” was the basis of Herrnstein's claim that if all inequality was removed people would sort themselves into “castes” that would be perpetuated from one generation to the next. Heritability is the proportion of variance in a trait in populations attributed to inherited, genetic factors. We have already seen that when alleles at several loci influence a trait, most people with the trait in successive generations will not come from parents or ancestors who also manifest the trait. No matter how high the heritability, it does not support Herrnstein's argument. For the same reasons, claims that the heritability of IQ is high, and that the average IQ of the lower social classes is less than more advantaged groups, cannot support the argument that the poor threaten the gene pool of the entire population.25 Furthermore, the measure of heritability in human populations tends to exaggerate the genetic component of variation.
Heritability is often measured by comparing variance of a trait among identical (monozygotic) and fraternal (dizygotic) twins and full, non-twin siblings. Full, non-twin siblings and fraternal twins share half of their alleles whereas identical twins share all of their alleles. In measuring heritability, it is assumed that the intrauterine and postnatal environments of non-twin siblings in a family will differ whereas the intrauterine environment of a fraternal or identical twin pair will be the same for both members of each pair and their postnatal environments will be equally similar (fig 1). Thus if a higher proportion of identical twins resemble each other in the trait being measured than do fraternal twins, then the greater the contribution of the genes to the trait and the higher the heritability. If, on the other hand, the proportion of identical twins who resemble each other in a trait is no greater than fraternal twins, and a higher proportion of fraternal twins show the trait than do non-twin siblings, then the greater the contribution of environment and the lower the heritability.
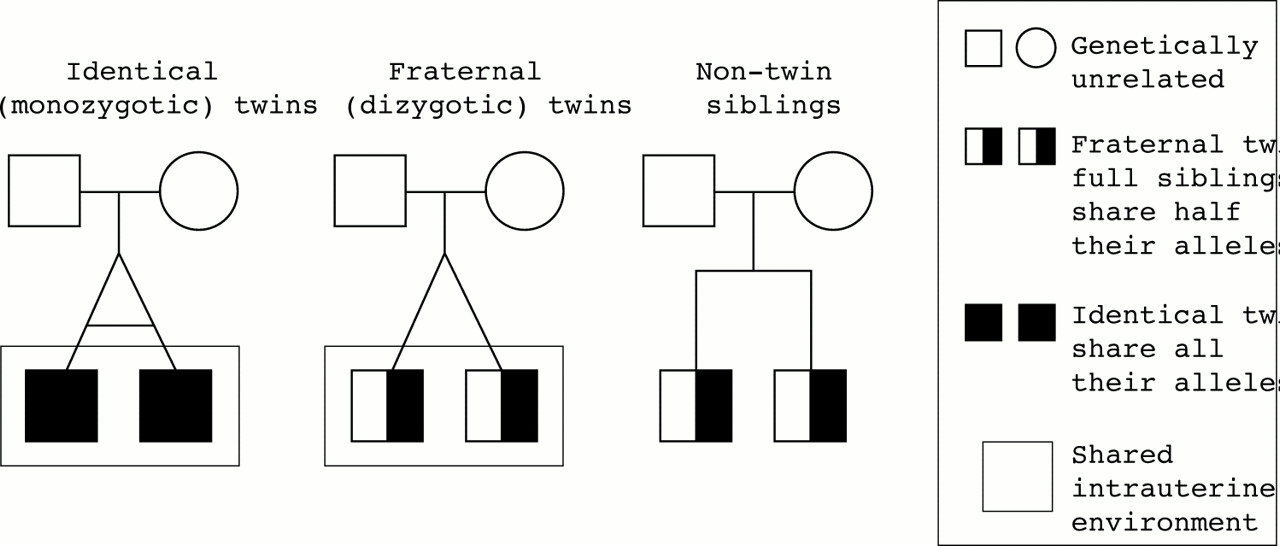
Comparison of identical, fraternal, and non-twin full siblings showing the environmental sharing of both types of twins assumed under measurements of heritability.
Harrap and Hopper recently pointed out that twin studies are biased towards higher estimates of heritability than the biology of twinning warrants:
“Twin studies depend on the assumption that the effects of environmental factors shared by twins are similar for dizygotic and monozygotic pairs. Yet few would also deny that lifestyle, diet, and behavior, and even intrauterine life are shared more closely between monozygotic than between dizygotic twins. Any greater shared environment in monozygotic pairs will be misattributed to additive genetic factors, thereby augmenting heritability estimates.”26
A more realistic portrayal of the environmental-genetic relations in different sibling pairs is shown in figure 2. (To carry out heritability measurements with statistical confidence, one needs large numbers of twin pairs and non-twin siblings. For simplicity sake, only one pair of each type of twin is shown in figures 1 and 2.) A large proportion of fraternal (dizygotic) twins develop in separate chorionic compartments and are, therefore, quite isolated from each other and exposed to different environments. (Even two identical twins may differ in their intrauterine environment; for instance, one twin may “bleed” into the other. This occurs in 10%–15% of monochorionic pregnancies.27 In the more accurate depiction of figure 2, the heritability estimate would be lower for any excess of sharing of a trait by identical than fraternal twins than would be the case if figure 1 held; it underestimates the environmental differences.

{kind=link}
{kind=link}
Comparison of identical, fraternal, and non-twin full siblings approximating the actual degree of environmental sharing of both types of twins.
As usually measured, heritability neglects genetic-environmental interactions (see for instance Lichtenstein et al 28). As these interactions will vary with the environment (holding the genetic component constant), the heritability of a trait in one environment will be different than in another.29 The heritability of reading disability would be much higher when no twins or siblings in the study were allowed to wear corrective eye glasses than if those with myopia were allowed to wear them. A recent study found that even rigorous attempts to maintain a constant environment of the same inbred (genetically identical) strain of mice did not succeed; mice differed in their behaviour, which could only be attributed to subtle environmental changes.30 It would be impossible to study the heritability of determinants of social class until the environments of all social classes are the same. And this would destroy the concept of social class. Allen has concluded:
“Short of breeding large numbers of couples and raising their children in highly controlled environments, which for moral and ethical reasons I hope we never contemplate, to try and separate hereditary from environmental components of human social behavior seems largely futile.”3
Measuring the extent of differences between identical twins reared apart and those reared together is also used to estimate the relative contributions of environment and inherited genetic factors. Separated identical twins that have been studied are quite similar to each other in many traits, arguing for high heritability, but the studies may be biased in favour of recruiting the most similar pairs.31 Moreover, separating human twins is not done as part of an experiment to study heritability but for some social necessity. Many of them live in similar environments, including the same village and the same school, and may be cared for by close relatives. In an analysis of one twin study, Kamin noted the intelligence quotient (IQ) correlation of 27 monozygotic twin pairs reared by relatives in separate households was 0.83, while for the 13 pairs raised in unrelated families it was 0.51, significantly lower31 (50). Kamin concluded, “this is very powerful evidence for the role of environment in determining intelligence scores.”
Observing the extent to which non-twin siblings placed for adoption early in life more closely resemble their biological than their adopting parents is also used to estimate the extent to which a trait is inherited. When IQ is used, the correlation between biological parents' and adopted siblings' is very high; there is a genetic effect. However, the average IQ of the adopted siblings is significantly higher than their biological parents and is closer to the IQs of their adopting parents. Again, adoption is not done as a social experiment. Lewontin comments:
“people who decide on an early adoption for their children are usually working-class or unemployed people who do not share in the education and culture of the middle class. People who adopt children, on the other hand, are usually middle-class and have an appropriate education and cultural experience for the content and intent of I.Q. tests .”32
In a study addressing this point, the IQ of children born to parents with less than nine years of school (group B-) who were adopted by parents with more than 12 years of school was 12 points higher than that of group B-children adopted by parents with less than 9 years of school.33
Heritability was resorted to when it became evident that the inheritance of complex traits did not follow the Mendelian ratios expected of traits determined by alleles at a single gene locus. This is not to say that the alleles at each gene locus contributing to a complex trait are not inherited in Mendelian fashion. Fisher showed many years ago that even continuously varying traits, like stature, span, and forearm length, can be explained by the inheritance of alleles at multiple gene loci in accord with Mendel's laws.34 Heritability measurements may be useful in humans in comparing the inherited genetic contributions to different diseases in approximately the same environments, as Lichtenstein et al recently did for cancer.28 Childs has pointed out that as environmental factors that increase the risk of disease (or of undesirable traits) are discovered and corrected, people who continue to display the disease (or trait) are more likely to have genetic susceptibilities; that is, over time with environmental interventions, heritability will increase as the proportion of the population with the trait decreases.35 Heritability says nothing about the number of gene loci that influence a trait nor, consequently, about how genes influence it. Neither, as we have seen, does it say anything about the perpetuation of a trait from one generation to the next.
SUMMARY AND CONCLUSIONS
It is no exaggeration to say that every gene will soon be identified and at least some of each one's functions elucidated. Both scientists and the mass media have, however, made exaggerated claims of the explanatory value of genes for complex diseases, and behavioural traits, including some that correlate with social status.36–38 An appraisal of genetic principles and of the function of genes, as I have attempted in this paper, should dispel claims that phenomena that are largely influenced by the environment and social interactions can be attributed to our genes. When they do play a part, the contribution of genes will be highly complex and difficult to unravel, involving the interactions of genes with each other and with a variety of environmental factors.
This complexity also makes it virtually impossible that the same genetic variants will be concentrated in any social class and transmitted more to children of that class than to the children of another class. The myth of inherited class differences is a justification for maintaining a ruling class in power, for limiting the ability of the lower classes to procreate, and for minimising social support programmes for the poor. This last is based on the erroneous belief that genetic factors (either for single gene or complex traits) cannot be compensated or corrected. The deleterious effects of several genetic diseases, including PKU, can be prevented.
Nor is there evidence that genes explain the alleged differences between so called races. Races differ strikingly only for alleles at very few gene loci, primarily those that affect response to climatic differences 23 (chapter 1). As with social class, the claim that race is based on genetics is used as an unwarranted justification for discrimination.
The roots of social class differences do not lie in our genes. By reducing inequities between classes and providing equal opportunity, we will not see “families sustaining their position on the social ladder from generation to generation,” as Herrnstein predicted. Applied correctly, genetic principles and methodologies will cease to provide a subterfuge for those seeking to justify the maintenance of social inequality.